







Митохондриите се срещат във всички растителни и животински клетки. Изкючение са едноклетъчните рукариотни организми, живеещи постоянно в анаеробна среда, дифинитивните еритроцити, в роговелите епителни клетки, мутантни дрожди, загубили изцяло или частично митохондрииния си геном. Митохондриите има собствена ДНК и се възпроизвеждат независимо от ядребата ДНК. Наблюденията на митохондриите датират от края на 20в като им са давани различни имена свързани с формата им (митозоми, хондриозоми, биобласти и да. ) В 1898 г. Бехда въвежда името митохондрии, а през 1900 г. Михаелис предлага използването на багрилото. Янус гриин Б, специфично багрещо митохондриите (хистохимичен маркер)
ФОРМА – варира силно дори в пределите на една клетка – специфичниелипсовидни, бъбрековидни, силно източени – нишководни.
РАЗМЕРИ – Дебилината на митохондриите е относително потоянна около 0,5 мкм. Дължината им се коледае от 0,5 до 7-10 мкм при хишководните (силно източените) форми.
Количеството на митохондриите в клетката е различно в зависимост от функцията на клетките.
БРОЯ на митохондриите в клетката зависи от нуждите на самата клетка от енергия и АТФ.
ЛОКАЛИЗАЦИЯТА на митохондриите също е различна, обикновено те се разполагат в тази част на клетката, където има нужда от енергия и АТФ. При недиферинциралихондриите са клетки митохондиите са разположени равномерно в цитоплазмата, а при диферинцираните като например мускулни, митохондриите се намират в близост миофибрилите; в сперматозоидите – завити са като спирала около камшичето и т.н.
СТРУКТУРА
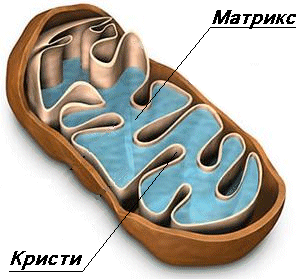
Класическите митохондрии се представят като елипсовидни органоиди, ограничени с две мембрани (двумембранни органоиди), които ги разделят на две пространства – интермембранно пространство и вътрешен матрикс.
Вътрешна митохондриална мембрана – дебелината около 7 nm – отделя съдържанието на органоида от цитоплазмата ( хиалоплазмата). Тя нормално има гладък контур и не е свързано с други мембрани в цитоплазмата притежава неспецифична проницаемост. По статична е, с белтъчно съдържание около 20%. Съдержа порови белтъци – порини през които могат да преминат всички молекули до 5000 D. Други белтъци в тази мембрана включват ензими, свързни с митохондриалния липиден синтез.
Интермрдиарното пространство е изпълнено с хомогенно течно вещвство със слаба електронна плътност. Съдържа предимно нискомолекулни съединения – кофактори и субстрати на биологичното окисление.
Вътрешна митохондриална мембрана – ограничава матрикса (митоплазмата). Тя образува многобройни вгъвания, наречени крисити. Тези нагъвания
(инвагинации) многократно увеличават повърхността на вътрешната мембрана. Криситите са насочени навътре към марикса Вътрешната мембрана се различава от външната. Тя е с висико специфинчна проницаемост, тя е по-гъвкава и по-динамична, с белтъчно съдържание 75%. Това е свързано с електрохимичния градиент на Н , който стимулира АТФ синтезата. Най-общо белтъците на вътрешната мембрана изпълняват три типа функции:
- Узвършват окислителноредукционните реакции в електрон-транспортната верига.
- Синтезират АТФ в матрикса, чрез АТФ – синтези в мембраните.
- Позволяват преминаването на метаболити навън и навътре от матрикса, чрез транспортни белтъци.
МАТРИСК
Висико концентрирана смес от стотици ензими, включително и ензими необходими за цикъла на Кребс. Матриксса има финнозърнест и хомогенен строеж. В него се наблюдават тънки нишки и гранули. Нишките са няколко копия на молекули ДНК а гранулите – митохондриални рибозоми. В матрикса се срещат по-големи гранули които са депо за отлагане на калциеви и магнезиеви соли.
МИТИХОНДРИАЛНА ДНК
Респликацията на митохондриалната ДНК протича независимо от репликацията на ядрената ДНК. Обикновенно репликацията на митохондриалната ДНК става след завършване репликацията на ядрената ДНК. Митохондриалната ДНК е двуверижна, за нея е характерна суперспирализация но има атсъствие на белтъци (при ядрената имаме хистолин). Тя обикновенно се прикрепва вътрешната митохондриална мембрана, т.е за митохондриите с характерна автономия при възпроизвеждането, но по отношение на пълна автономност относно синтезата на пълния набор от белтъци необходими за изпълнение на изпълнение на функиите си, митохондриите често използват белтъци синтезирани от цитоплазмените рибозоми, които постъпват в матрикса и се локализират на специфичните на тях
места.
ФУНКЦИИ НА МИТОХОНДРИИТЕ
- В тях се извършва цикъла на Кребс
- Окислително-фосфолиране ( синтез на АТФ )
- Бета – окисление на мастни киселини
- Елонгиране ( удилжаване ) на мастни киселини
- В митохондриите се отстраняват някой токсични събстанции.
Митохондриите са сързани главно с енергитичния метаболизъм на еукариотната клетка клетката – синтез на АТФ. Стратегията на цикъла на Кребс е същата с тази на гликозата ( извършва не при анаеробни условия в цитоплазмата),а именно: Разграждане на голени въглеводородни молекули с отделяне на енергия по формата на АТФ или под формата на НАДН2 и ФАДН2
ОКИСЛИТЕЛНО – ФОСФОРИЛИРАНЕ – Водорода от НАДН2 и ФАДН2 се подава по веригата от най-малко пет междинни субстанции [Тази верига се нарича електрон-транспортна верига (дихателна верига)]. Този водород по тази верига достига до молекулата на О2 и се образува Н2О. Всяко пренасяне на Н редуцира (приема е-) на донорната молекула.
Преноса на Н в началото на веригата, а в края на е- създават електроден градиент който е причина за фосфолиринато на АДФ до АТФ. В този процес участват оксизолите в чиито главички е разположен АТФ – цинтетазния комплекс
Я-ОКИСЛЕНИЕ – Свободни мастни киселини се отркиват в клетката само в следи, защото са силно токсични. В цитоплазмата масните кислелини се свързват с глицерол в моноди и триглицериди и фосфолипиди. Те се хидлолизират в цитокл. и се транспортират в митохондриите кдето се окисляват напълно
(Я-ОКИСЛЕНИЕ )
ЕЛОНГОРАНЕ НА МАСТНИ КИСЕЛИНИ – Най-общо мастните киселини се синтезират в агранулирания едноплазмен ретикулум но в митохондриите също има ензими катализаращи елонгирането на палмитинова и др. наситени мастни киселини.