








Ако сте решили да плувате или се гмуркате във водите на Австралийските северни територии, трябва да знаете, че това може да е опасно за живота. Едно слабо опарване от почти незабележима полупрозрачна малка медуза, може да Ви докара големи главоболия. В началото почти няма да усетите нищо, а може и да не забележите кога сте били опарени, но след около 20-30 минути, ще получите изключителни болки по тялото, най-вече в гърба, главоболие, втрисане, гадене, повръщане, мускулни крампи по краката и корема, силно повишаване на кръвното налягане, което вторично да доведе белодробна едема, мозъчен кръвоизлив (инсулт) и смърт. Болките са толкова силни, че дори и морфин не помага. Това състояние продължава дни, а в някои случаи до две седмици. Описано е като Ируканджи синдром и е доказано, че се предизвиква от допира до малките кубомедузи, наречени още Ируканджи медузи.
Името Ируканджи ( Irukandji) е взето от английската версия на името на аборигени, пазители на земята, между Кернс и порт Дъглас (the Yirrganydji people), където синдрома Ируканджи е за първи път регистриран. За първи път този термин се използва през 1952 г, когато са регистрирани случаи на тежки последствия от едно малко ужилване/опарване, без да има видими наранявания на кожата. Сауфкот и Поуис през 1944, наричат това явление „ужилване/опарване тип А”. Късната 1944 г, един непознат военен доктор в Нова Гвинея има подобен случай, но той регистрира кожни лезии около мястото на опарването и агент, който ги е причинил /малка прозрачна медуза/. Преди този случай има спорадични съобщения за подобни такива по света. През 1961 г, доктор Джак Барнс от Кернс доказва, че все още некласифициран вид медуза предизвиква Ируканджи ефект. Той използва индивид, за да опари себе си, 9 годишния си син и доброволец, спасител на сърфисти. Тримата оживяват след последствията.Тази медуза по-късно е наречена Carukia barnesi, Карукия на Барнс, в негова чест. Само 40 години по-късно, към групата на Ируканджи медузите се добавят и други видове, които причиняват такива симтоми, макар и с вариации.
История на изследванията на Ируканджи синдром
1935-1936 На Медицинска конференция в Кернс, се препоръчва запознаването със събрана информация за опарвания/ужилвания, включително от морски животни, в околността на Северен Куинсленд. Това е направено под егидата на доктор Хюго Флекер, който започва да регистрира такива случаи от началото на 1935, включително и тези, които по-късно са разпознати като Ируканджи синдром
1944 – През 1964 г., ретроспективно са обявени два случая от 1944г на Ируканджи синдром, на остров Нумфор, северозападно от Папуа. Описано е, че това е предизвикано от малка /3-5 см/, почти безцветна медуза. Информацията е от местен шаман/доктор.
1945 – Първото официално обявление, на основа на симптоматиката и епидемиологията на това, което сега се знае като Ируканджи синдром, групирани като категория на „поражения от неизвестен агент, върху къпещите се в северен Куинсленд”.
1952 – Сауфкот ретроспективно обявява около 90 случая на „остър, засягащ цялото тяло ефект, без видими обриви” от плажовете около Кернс през лятото на 1943 – 1944 г и го нарича „опарване тип А”
Терминът „Ируканджи синдром” за пръв път се използва като описание на случаите около района на Кернс, по името на местните пазители. Флекер започва да определя епидемиологията на този вид опарвания, базирайки се на случаи от дългогодишен регистър на Северен Куйнсленд.
1960 – Барнс детайлизира серия от Ируканджи опарвания, включително сравнителни данни от такива случаи около Кернс, регистрирани случаи от Амбулаторния център в Кернс и Базовата болница, за периода от 1956 – 1960г.
1961 – На 10 декември, Барнс хваща малка медуза и експериментално опарва себе си, сина си и един доброволец, за да може да потвърди версията си за агента, предизвикващ Ируканджи синдром
1967 – Медузата, предизвикваща Ируканджи синдром е описана и класифицирана като Carukia barnesi
1970 – Пусната е за ползване първата противоотрова за Ируканджи синдрома / CSL Ltd /. Барнс докладва, че за съжаление тази противоотрова е неефективна.
1986 – Първото официално съобщение, че противоотрова за кубомедузата Chironex fleckeri, е използвана за случаи на Ируканджи синдром, но без особен ефект.
Предполага се, че α–и β– адренорецепторни антагонисти могат да се използват за противодействие на проявите на Ируканджи синдрома и специално по отношение на повишаването на кръвното налягане.
1987 –Остър белодробен едем, с дисфункция на лявата сърдечна камера, е определен като част от Ируканджи синдром.
1988 – Барбара Кинси съобщава за определянето на различни видове кубомедузи, предизвикващи подобен на Ируканджи синдрома, с вариации при отравянето
1997 – Съобщение за първата жертва на Ируканджи синдрома, предизвикан от опарване на кубомедуза.
Първо съобщение за папилоедем и кома, свързани с Ируканджи синдром, дължащи се вероятно на церебрален едем.
1998 – Случай с подобен на Ируканджи синдром в Геелонг Болница във Виктория
2000 – Публикувано е, че екстракт от Carukia barnesi предизвиква масово освобождаване на катехоламини в опитни животни.
Съобщение, че животозастрашаваща сърдечна недостатъчност се наблюдава при Ируканджи синдром
2000 – Към Университета Джеймс Кук в Кернс се открива център TASRU – Tropical Australian Stinger Research Unit, чиято работа се фокусира върху биологията на кубомедузите и медицински направления при опарване с тях.
2001 –Съобщение за подобен на Ируканджи синдром случай от Хавай, САЩ
В Куинсленд са регистрирани два смъртни случая, вследствие на Ируканджи, като смъртта е настъпила от интрацеребрален кръвоизлив, предизвикан от високото кръвно налягане.
2002 При остъргване на мястото на опарване е идентифициран нематоцист, който не принадлежи на Carukia barnesi, така се потвърждава хипотезата, че Ируканджи се предизвиква не само от един вид медуза. Тези нематоцисти са определени на нов Ируканджи вид – Malo kingi. По-късно се съобщава, че нематоцистите принадлежат на Carukia barnesi, но са различни от обичайните.
Ируканджи синдром е регистриран във Флорида , САЩ
2003 – Интравенозно вливане на магнезии за пръв път е използвано за лечение на последствията от Ируканджи синдрома.
2004 – Съобщава се, че интравенозния магнезий се използва ефективно при лечение на болката и високото кръвно налягане от Ируканджи ефекта
2004 – Документиран е Ируканджи ефект в Брум, Австралия.
2005 – Описани са нови родове и видове Ируканджи медузи: Malo maxima; Carukia shinju; Alatina mordens; Gerongia rifkinae
Фармакологичен анализ на екстракт от отровата на Carukia barnesi , потвърждава освобождаването на катехоламини и модулацията на невралните натриеви канали.
Регистриран е Ируканджи подобен синдром в Гваделупе, Кариби.
2007 – Потвърден е нов вид Ируканджи медуза – Malo kingi и с нея е свързан фатален случай от 2002 г.
2008 – Определен и класифициран още един вид Ируканджи медуза – Morbakka feneri
2008 – Експериментално е демонстрирано освобождаване на катехоламини предизвикано на отровата от Alatina mordens
2009 – Непубликувана десертация, съобщава за създаването на първите клонални ДНК библиотеки на Ируканджи медузи.
Съобщения за случаи на Ируканджи синдром за период от 1997 до 2007 в Северната територия, Австралия
2010 – Съобщение за Ируканджи синдром случай в Малайзия.
2011 – Публикувани са данни за експериментални изследвания, които потвърждават подобен на Ируканджи синдром, ефект върху симпатиковото нервнотрансмитерно освобождаване от отровата на Malo maxima
2012 – Напълно е секвениран митохондриалния геном на Alatina moseri
CSIRO разработва първоначален модел на прогнозиране на климатични условия, с цел да се предскаже масова поява на Ируканджи медузи.
http://www.marinestingers.com.au/index.cfm
http://www.jcu.edu.au/stingers/index.htm

Кой предизвиква Ируканджи синдром?
Живеещите по крайбрежието кубомедузи са малък клас книдарии, наречен Cubozoa, който е близък със същинските медузи Scyphozoa. В клас Cubozoa досега са описани досега 50 вида, в 7 семейства, които се отличават с няколко забележителни качества: наличие на очи, свързани с визуални способности; необичайно ухажване и брачно поведение; изключителна токсичност. Тези техни интригуващи качества ги поставят в центъра на научен и публичен интерес. Жизнения цикъл на кубомедузите се състои от полипна фаза – прикрепена и възрастна подвижна фаза – медуза, подобно на същинските медузи. Слабото разнообразие в рамките на класа се обяснява или с еволюционна история /липсата на алопатрично видообразуване/ или с недостатъчно таксономична изучаване на групата. Cubozoa се дели на два монофилетични разреда Chirodropida и Caribdeida. Името на медузите идва от формата на тялото, което е по скоро кубично или с формата на камбанка /чадърче/, като долната част има капак наречен велариум, с помощта на който се концентрира и увеличава потока вода, с цел движение. Нервната им система е много добре развита, като в основата на чадъра имат нервен пръстен, който координира движенията. Обикновено пипалата надвишават неколкократно камбанката, и именно там са разположени парливите клетки – нематоцистите.
Разпространени са в тропиците и субтропиците, като някой достигат и до умерените ширини. Ката цяло видовете не са добре описани и изследвани по отношение на екология, систематика, токсичност, етология, физиология, генетика и т.н.
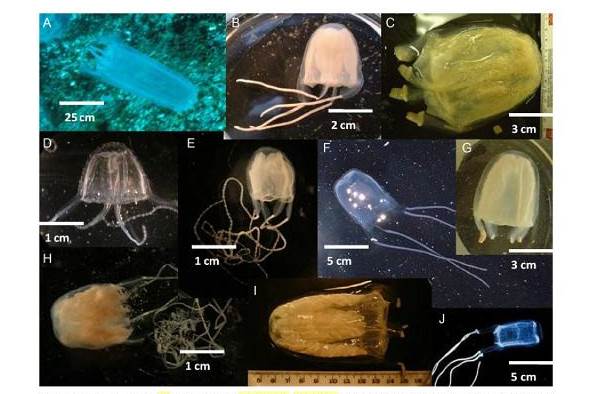
Фиг. 1 Видове Австралийски Ируканджи медузи: /А/ Alatina sp. От Нингалу риф, Западна Австралия; /В/ Malo kingi от Северен Куинсленд; /C/ Morbakka fenneri от Централен Куинсленд; / D/ Carukia barnesi от Северен Куинсленд; / E/ Carukia shinju от Брум, Западна Австралия; / F/ Malo maxima от Брум, Западна Австралия; / G/ Gerongia rifkinae от Северните Територии; /H/ Carukia sp. От Големия Бариерен Риф; / I/ Alatina mordens от Големия Бариерен Риф; / J/ Morbakka sp. От Ню Саут Уелс
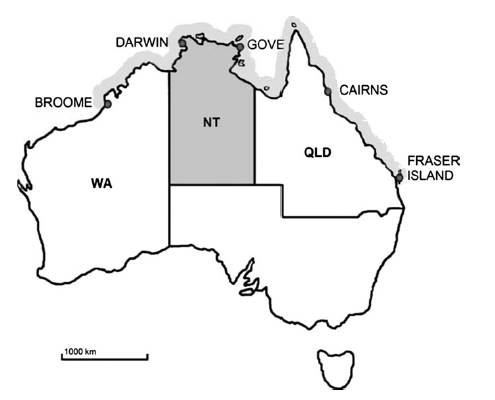
Фиг. 2 Географско разпространение на Ируканджи синдрома в Австралия. NT – Северни територии, QLD – Куинсленд, WA – Западна Австралия. Сивите участъци на крайбрежията показват географското разпространение на синдрома Ируканджи.
Еволюционната история на Cubozoa не е добре изследвана, но вследствие от изясняване на филогенията, основано на анализ на рибозомни, нуклеарни и митохондриални гени са направени няколко хипотези по отношение на еволюцията на токсичността и биогеографията. Първо, че последния най-близък предшественик на съвременните видове е притежавал способността да предизвиква Ируканджи синдром. Второ, основното разделение на Атлантически и Индо – Тихоокеански клонове може да се обясни с изолация, вследствие на древно събитие. Трето, счита се че сексуалния диморфизъм е възникнал еднократно, заедно с комплексно брачно поведение.
Важно откритие, вследствие от филогеографския анализ е, че видът Alatina moseri от Хавай и Alatina mordens от Австралия, са всъщност един и същ вид, представен с една непрекъсната метапопулация, разширила се посредством природни и/или антропогенни фактори.
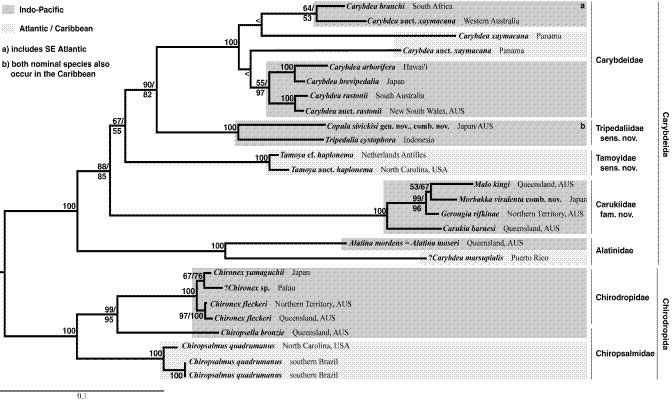
Фиг. 3 Филогенетични отношения, основани на данни от изследвания на митохондриална и нуклерна ДНК на кубомедузи
Еволюцията на токсичността на кубомедузите представлява сериозен интерес, тъй като е известно, че основна част от тези медузи са силно токсични, в резултат предизвикват големи загуби, както в сферата на общественото здраве, така и на туристическата индустрия, най-вече в Австралия. Усилията положени за изолиране на компонентите на отровата и създаването на антиотрова, по-специално за смъртоносната Chironex fleeckeri , като цяло не са успешни. В тази връзка, филогенетични изследвания на токсичността на видовете би била в помощ, още повече че е логично родствените видове да проявяват сродна токсичност. Токсичността варира между отделните видове, като при едни тя е безвредна за хората, а при други може да убие човек за минути. Chironex fleeckeri от Chirodropida е най-отровната медуза, позната досега. Секвенирането на хемолитичните протеини от отровата на две карибдеиди и три хиродропиди показват умерена дивергенция. Въпреки ,че в рамките на Cubozoa между аминокиселинните последователности на токсичните протеини са наблюдавани различия, модели на вторичната структура предполагат най-малко два общи структурни мотива, които са свързани с цитолитичната активност. Няма открит хомоложен протеин извън групата на кубомедузите и затова се счита, че те притежават ново и уникално семейство протеини.
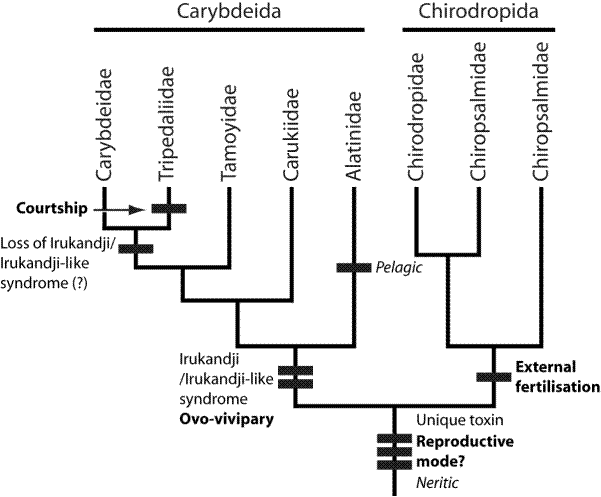
Фиг. 4 Проследяване на токсичността при филогенетичните отношения на кубомедузите
Ируканджи синдрома, предизвикван от отровата на няколко вида кубомедузи, първоначално се смята, че е характерен само за един вид – Карукия на Барнс, Carukia barnesi, но впоследствие се описва и за други видове – Morbakka, Alatina, Malo, Gerongia. Обикновено Ируканджи синдромът на другите видове, се определя като подобен на него, тъй като има същите базови симтоми на класическия синдром, предизвикван от Карукията на Барнс, но те могат да бъдат не толкова остри. Всички видове кубомедузи, предизвикващи Ируканджи синдром, спадат към Caribdeida. Сем. Carukiidae включва родовете Carukia, Morbakka, Malo, Gerongia. Най-рано се отделя в еволюционен план Alatinidae, след това Carukiidae и Tamoyidae, а последни Carybdeidae и Tripedaliidaе. Като за последните се предполага, че губят токсичността. Тази топология позволява да се предположи, че общия предшественик е притежавал възможността да предизвиква Ируканджи синдром.
Въпреки изясняването на филогенията, съществуват няколко проблема при изследването на токсина на кубомедузите. Трудно се изолира отровата, като се използват цели тентакли, вместо отделни нематоцисти и има доста противоречиви резултати между отделните изследователски групи. Токсините получени от различни части на една медуза и от различни индивиди на различен онтогенетичен стадий, се различават, както се различават и тези получени от отделни индивиди на един вид, но от различни географски региони.
Биогеографията на кубомедузите не е добре документирана по средни географски скали /провинции, региони или държави/, поради липса на данни. На голяма географска скала /океани/ има по-голяма яснота. Голяма дивергенция се открива между Индо – тихоокеанските и Атлантическите клонове. При Chirodropida, рода Chiropsalmus е изключително Атлантически, докато Chiropsella и Chironex са към Индо – Тихоокеанската група. Подобно е разделението и при Caribdeida, където Tamoyidae е ограничен в Атлантика, а видовете от Carukiidae са известни от Австралия и Япония, но е много вероятно да са разпространени и в целия Индо – Тихоокеански регион.
Друга характерна особеност, която най- вероятно ще се потвърди за всички видове кубомедузи е, че те не пресичат местообитанията през океана. Обикновенно се придържат към шелфа на даден регион. Така разделението на два основни клона – Индо – Тихоокеански и Атлантически, може да се обясни с древно събитие, което ги е разделило и изолирало. За съжаление фосили от медузи са голяма рядкост и по тази причина трудно би се установило с точност кога и какво се е случило.
Определени са няколко таксономични ключа за определяне на различните видове кубомедузи от Bentlage et al. (2010)
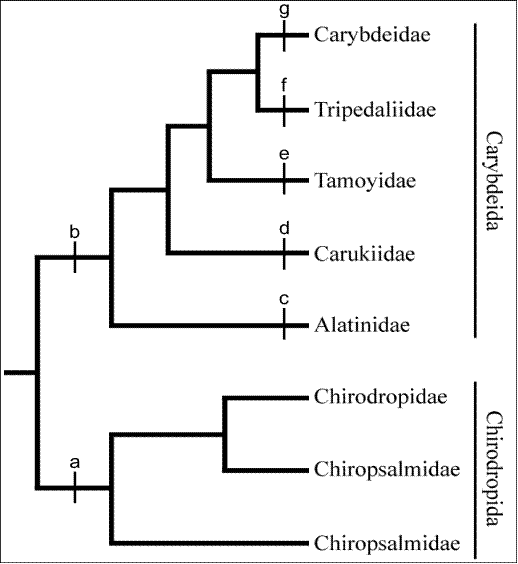
Фиг. 5 Родствени връзки при кубомедузите, съгласно посочените от Bentlage et al.(2010) таксономични ключове, посочени на кладограмата, разделящи разредите и семействата. Chirodropida имат няколко тантакли на педалиум /а/, докато Carybdeida имат само по един / b/. При Tripedaliidae, всеки педалий, носи по един тентакъл, но създава впечатлението, че има много тентакли на един общ педалиум. Семействата от Carybdeida се идентифицират по следните характеристики: Alatinidae /с/ – остиума на ропалийната ниша е т – образен; Carukiidae /d/ – остиума на ропалийната ниша с ропалиални рогца, стомаха няма гастрофоцели; Tamoyidae /е/ – остиума на ропалийната ниша е с форма на намръщена устна, липсват ропалийни рогца, гастрофоцелите са вертикални; Tripedaliidae /f / – остиума на ропалийната ниша е с форма на отвор на ключалка или по изключение като намръщена устна, има по 2 – 3 педалия на ъгъл; Carybdeidae /g/ – отвора на ропалийната ниша е сърцевиден.
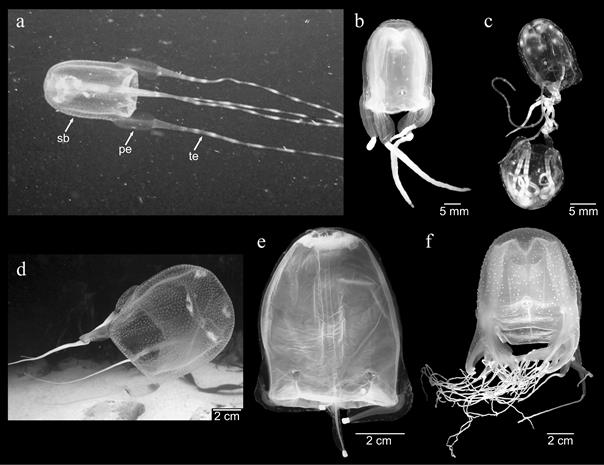
Фиг. 6 Представители на кубомедузи от всяко семейство на Carybdeida (a-e) и една форма от Chirodropida (f). Вижда се, че последните имат много тентакли на педалиум. a) Tamoya (Tamoyidae); b) Malo kingi (Carukiidae); c) мъжки и женски на Copula Sivickisi, копулирайки (Tripedaliidae); d)Carybdea branchi (Carybdeidae); e) Alatina sp. (Alatinidae); f)Chiropsalmus quadrumanus (Chiropsalmidae). sb: камбанката, куба на кубомедузите; pe: педалиум; te: тентакъл
Важен и характерен белег за медузите са техните нематоцисти или копривни / жилещи/ клетки. Нематоцистите преставляват сами по себе си капсула с намотан „ харпун”, потопен в отрова, със спусък /косъм / в единия край. Бързото му освобождаване, води до експлозивно изстрелване на „ харпуна”. Когато е освободен, нематоциста се поделя на три части: луковична капсула, стрела за пробиване, дълга подвижна тубула, която носи голяма част от отровата. Идентификацията на нематоцистите се основава на големината и формата на капсулата, броя и разположението на иглите на стрелата. Ируканджи медузите имат различни типове и размери нематоцисти, които се използват за видово определяне и характеристика на ужилванията. За характеристика на опарването, нематоцистите се получават или чрез остъргване от кожата или чрез стикер – проба, като последната е за предпочитане, тъй като не плаши пациента и не разрушава нематоцистите. При някой видове нематоцистите са различни типове, освен това се променят онтогенетично. Пропорциите на даден тип немотицисти спрямо друг тип, също се използва като характеристика. По принцип нематоцистите са разположени на тентаклите във вид на ивици /бандове/ , но при някой видове има и на камбанката. Бандовете нематоцисти са видовоспецифични и често се използват при определяне на видовете.
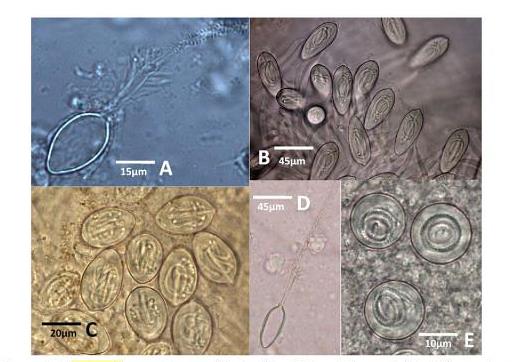 Фиг. 7 Ируканджи нематоцисти. /А/ Carukia shinju, освободен нематоцист; / B/ Malo maxima, неосвободен; /C/ Carukia shinju, неосвободен нематоцист / D/ Malo kingi, освободен; /E/ Carukia shinju.
Фиг. 7 Ируканджи нематоцисти. /А/ Carukia shinju, освободен нематоцист; / B/ Malo maxima, неосвободен; /C/ Carukia shinju, неосвободен нематоцист / D/ Malo kingi, освободен; /E/ Carukia shinju.
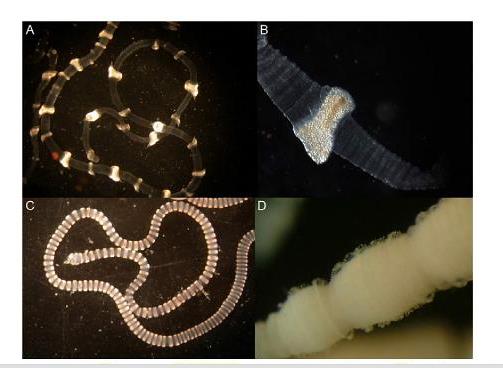 Фиг. 8 Типове тентакли при Ируканджи медузите. / A/ Carukia spp.; /B/ Carukia, отблизо; /C/ Caribdea, Malo, Gerongia, Morbakka, Alatina; /D/ Malo kingi, отблизо.
Фиг. 8 Типове тентакли при Ируканджи медузите. / A/ Carukia spp.; /B/ Carukia, отблизо; /C/ Caribdea, Malo, Gerongia, Morbakka, Alatina; /D/ Malo kingi, отблизо.
Отровите и токсините присъстват при не малко животни, не само за да улавят плячка, но и за собствена защита от други животни, а също така при седентарните форми за превенция на инфекции и болести. Тъй като мешестите успешно се развиват при условията на високо конкурентни хабитати от милиони години, не е изненадващо откритието, че притежават различни клетъчни изобретения и иновации. Биоактивни компоненти са открити при всички класове мешести и то в целия организъм, не изключително свързани с положението на нематоцистите. Химичния арсенал на Ируканджи медузите включва сложна смес от биологично активни вещества, включващ цитолизини, невротоксини и липази, пептидази, протеинови инхибитори и антимикробни вещества.
Друга характерна особеност на Ируканджи медузите са статолитите или баланс – камъчета. Всеки ропалиум има един голям статолит под клъстера с очите. Статолита расте посредством секреция на дневни растежни пръстени, подобно на отолитите на рибите или годишните пръстени на дърветата. Тези растежни пръстени на статолитите при Ируканджи, се използват за определяне на възрастта, а химичния състав може да покаже къде е живяла медузата. Формата на статолитите е родовоспецифична и служи за таксономичен белег. Тъй като са единствената твърда част, на иначе мекото тяло на медузите, статолитите са важни при идентификацията на фрагментарни проби, проби съхранявани в етанол или замразени, както и при фосили.
Кубомедузите имат комплексен жизнен цикъл, състоящ се от етап на първоначален, прикрепен, бентосен полип; етап на бавноподвижен полип и след почти пълна метаморфоза, етап на подвижна ювенилна медуза; накрая зряла разделнополова възрастна медуза. Точния жизнен цикъл и данни за метаморфозата липсват при повечето Ируканджи медузи.
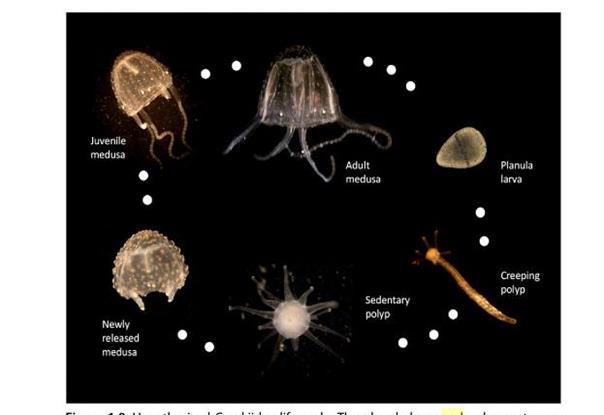 Фиг. 9 Хипотетичен жизнен цикъл на Carukiidae
Фиг. 9 Хипотетичен жизнен цикъл на Carukiidae
Най-интригуващата характеристика на кубомедузите е наличието на добре развити очи, въпреки липсата на относително сложно развит мозък. По това свое качество, да притежават комплекс от очи, кубомедузите са изключителни сред медузите, дори и сред безгръбначните въобще. Дълги години се дебатира, дали тези очи имат физическата възможност да разпознават изображения. Сега е ясно, че кубомедузите използват очите си да навигират, да откриват плячка и да се чифтосват. По-неясно е дали тези очи формират образи по начин, който ние все още не разбираме или се справят с размазани и неясни картини. По принцип кубомедузите имат 24 очи, групирани в 4 групи на всеки ропалиум. На средната линия на ропалиума има две комплексни очи, всяко с леща, ретина и роговица. Отстрани на тези очи има два чифта прости с пигментирани чашки, очи – оцели: един чифт цепнатовидни очи и един чифт ямковидни. Четирите групи очи имат различна структура и функция. Двете медианни очи са тип камера, със сферична или елипсовидна, подобна на тази при рибите леща, ретина и роговица. Има около 11 000 сензорни клетки във всяко око. Ретината е съставена от четири слоя – сензорен слой, пигментен слой, нуклеарен слой и слой на нервните фибри. Лещата е отделена от ретината с малко камерно пространство. Пигментния слой покрива отвън ретината, формирайки ирис около лещата. Зеницата на долното око може да се променя, съобразно светлинния интензитет, като променя отвора си за по-малко от минута, а на горното око е неподвижна. Горното лещовидно око е насочено нагоре, независимо от позицията на медузата и е установено, че се използва за сухоземна навигация.
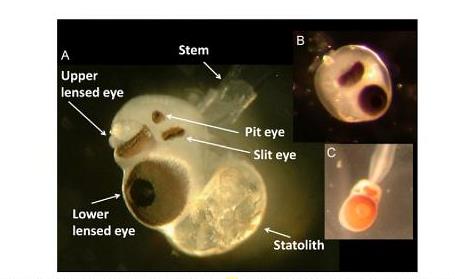
Фиг. 10 Очи на кубомедуза. – А/ Горно лещовидно око; Долно лещовидно око; цепнатовидно и ямковидно око; статолит от Chiropsalmus. В/ Липса на цепнатовидни и ямковидно око при Malo. С/ Изключително голямо долно лещовидно око при Alatina.
Долното лещовидно око е ориентирано надолу и е с малко по-широк диапазон на видимост. Страничните, цепнатовидното и ямковидното, очи имат различни структурни характеристики. Ямковидното око има единствено пигментирани фоторецептори и се счита, че служи за отчитане на осветеността без пространствена резолюция. Цепнатовидното око е съставено от четири типа клетки, включително кануподобна група на стъкловидни клетки, формиращи лещоподобна структура върху ретина. Това стъкловидна група има набраздена повърхност и се смята, че действа като УВ филтър. Предполага се, че това око има пространствена резолюция и служи за определяне на вертикалното движение.
Експериментални изследвания показват, че комплексните очи на кубомедузите могат да формират изображения. Кубомедузите са способни да отговарят на различни сигнали от заобикалящата ги среда, такива като форми, фигури, цветове на светлината и да реагират спрямо тях, обикновено предсказуемо. По-сложно поведение, такова като преследване, избягване, навигиране, ухажване и копулиране, включва използването на визуалните им способности и някои познавателно – поведенчески прояви.
Визуалните способности на кубомедузите определят една от основните поведенчески реакции – фототаксиса. Фототаксис е ориентацията им и придвижването по посока на светлината. Барнс е уловил своята Карукия именно насочвайки светлина към водата и е уловил привлечения от нея екземпляр. През деня, насочването на светлина не се отразява така, както когато се насочи на сумрак ,тогава се вижда ясно как кубомедузите са привлечени от нея. Именно тази им особеност, ги прави лесни за улавяне по този начин. Привличането им от светлината е важна особеност, от гледна точка на предпазните мерки, необходими да се вземат при работа на тъмно, когато се осветява само част от водата при риболов, водни строежи, пристанищни и яхтени съоръжения, както и при любителско гмуркане и други туристически забавления.
Един от основните въпроси, свързани с Ируканджи медузите е опазването на човешкото здраве. В тази връзка се изясняват разпространението на Ируканджи синдрома, биология и екологията на Ируканджи медузите. Редица учени считат, че проблема със този синдром е донякъде подценяван, защото се отчитат само регистрираните и разпознати като такива случаи, а е възможно те да са много повече, но първо, да не са разпознати проявите на синдрома и съответно да не е регистриран като такъв в болничните заведения; второ при смъртен случай, тъй като еритемата от опарване на Ируканджи медуза е сравнително слаба, напълно е възможно тя да е отшумяла post mortem и съответно случая да не е регистриран като причинен от Ируканджи медуза. Редица институции са поели ангажимент да разпространяват информация за характерните особености на медузите и синдрома, който предизвикват, както и насоки за първа помощ в този случай. Прави се статистика на регистрираните случаи по света с цел създаването на обща база данни.
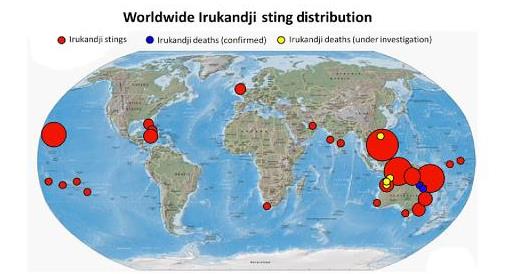
Фиг. 11 Разпространение на ужилвания от Ируканджи медузи по света. Големината на кръговете показва количеството на ужилванията за дадения регион. С червен цвят са регистрираните ужилвания; със син цвят потвърдени два смъртни случая; с жълт цвят непотвърдени четири смъртни случая.
Основния регион, за който Ируканджи медузите и съответно синдрома, са характерни е региона на Австралия.
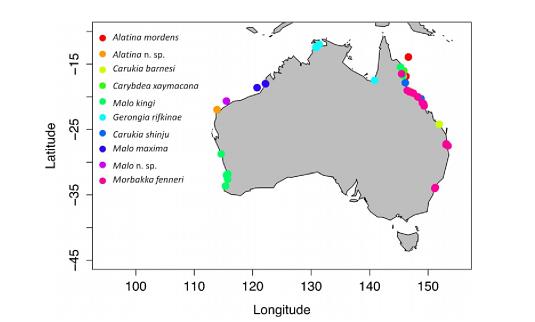
Фиг. 12 Разпространение на Ируканджи медузи в Австралия: потвърдените места са оцветени със съответния цвят.
Всички региони, където са регистрирани Ируканджи медузи се характеризират с това, че представляват олиготрофни шелфови хабитати. Обикновено са богати на водорасли и различно обрасли с тях, както са и характерни честите цъфтежи на салпите, Клас Thaliacea. Обикновенно местата с най-чести случаи на Ируканджи са свързани с цъфтежи или поне сериозно присъствие на салпи.
Вертикалното разпространение на кубомедузите по принцип се определя от осветеността. Като цяло те са описани, че се придържат към повърхността. Случаите на ужилване от тези медузи основно са по тялото или ръцете по време на плуване или стоене прав във водата. При анализ на данните на опарване по тялото, се прави извод, че най-вероятна дълбочина до която достигат тези медузи във водния слой е 50 см.
При анализ на събраните данни на 1629 ужилвания на кубомедузите в Австралия, за периода от януари 1893 до юни 2013, са показани статистически резултати кога това се случва през деня и в различните сезони.
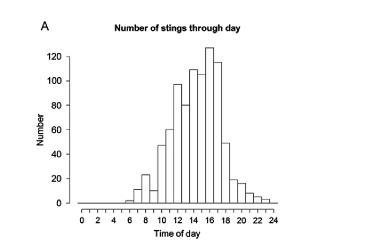
Фиг. 13 Ужилване от Ируканджи медузи в Австралия: А/ през деня и сезонност на източното крайбрежие / В/ и на западното крайбрежие /С/.

От дългогодишни наблюдения и от информация на спасители на плажа, става ясно че като цяло ужилванията на Ируканджи медузи не са постоянни и целогодишни, има сезонност, освен това тяхната поява и зачестяване се свързва с поява на специфични климатични промени, ветрове, течения и т.н
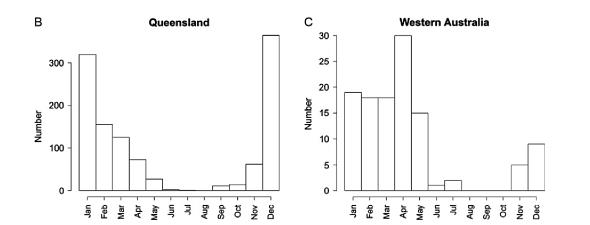
Местоположението на ужилванията също статистически показва, че най-голям брой от тези случаи са регистрирани за района на плажната ивица и то през лятото. В тази връзка от голямо значение е информираността и вземането на предпазни мерки от страна на туристите.
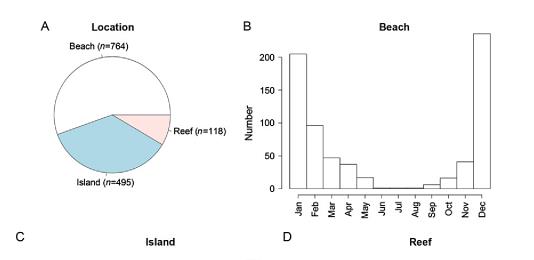
Фиг. 14 Ужилвания на Ируканджи медузи в Куинсленд по местоположение и сезон.
Наблюдения в Брум, показват, че ужилванията на Ируканджи медузи имат два пика и те се свързват с пиковете на мусоните, което още не е добре изяснено.

Друга особеност е способността им да формират репродуктивни струпвания. Дългогодишни наблюдения свързват появата им с лунарна периодичност. Например Alatina moseri формира такова репродуктивно струпване на 10-12 ден всеки месец, след пълнолуние и съответно немалко видове се изхвърлят на брега. Тази лунарна периодичност има малки сезонни вариации, но големи годишни такива. Например през 2001 г, са преброени повече от 10 000 екземпляра на такова струпване, докато през 2005 едва една четвърт от това.
Carukia barnesi образува цъфтежи с големи струпвания, докато Morbakka и Malo са описани като живеещи по- случаи на струпване и на двата вида по западните крайбрежия на Австралия. Тогава са били изхвърляни стотици индивиди по плажовете.
Основните усилия върху изучаването на екологията и биологията на Ируканджи медузите е насочена към установяване на зависимости, които да дават възможността да се предскаже появата им по плажовете и по този начин да се намали риска от ужилвания и проява на Ируканджи синдрома. За главни биоиндикатори се посочват салпите, които неизменно присъстват в големи количества по плажовете, когато са регистрирани случаи на ужилвания. Други индикатори са бризовете, духащи към континента и последващо затишие.
Най-ефективното предпазно средство срещу опарванията от Ируканджи медузи е цял предпазен костюм, изработен от ликра. От изпитваните различни видове материи, за създаването на такъв предпазен костюм, най-ефикасна се е оказала тази ликра.
За съжаление все още няма разработена ефикасна противоотрова и ваксина за Ируканджи синдрома. Като най-ефикасно средство за третиране на местата на опарване и намаляване силата на синдрома е обработване на мястото с оцет. В помощ за намаляване на симптомите на синдрома, доказано се използва венозен магнезиев сулфат. Той остава засега най-ефикасното средство за лечение. Обикновено в болниците третират отделните симтоми, засега няма средство, което да повлиява синдрома като цяло.

Автори:
Димитра Лефтерова
д-р Чавдар Черников
Източник: nauka.bg





