








Дезоксирибонуклеиновата киселина (ДНК) е открита през 1929 г. от Фебъс Арън Теодор Левен (1869 -1940), руски учен, работещ в Америка. Уникалната двойна спирална структура на молекулите на ДНК е описана през 1953 г. в Англия като резултат на обединените усилия на два екипа от изследователи. През тази година Франсис Крик (роден 1918) и Джеймс Уотсън от Америка (роден 1928), работещи в Кембридж, създават първия модел на молекулярната структура на ДНК, който отговаря на всички експериментални характеристики на субстанцията. Тяхната работа става възможна благодарение на откритията и идеите на Розалин Франклин (1920 – 1958) и Морис Уилкинс от Нова Зеландия (роден 1916), които работят заедно близо до Лондон и изучават молекулата на ДНК. През 1962 г. тримата, които са останали живи, си поделят Нобеловата награда за това откритие. Преди да е намерена неговата структура, вече са били разкрити наследствените функции на ДНК.
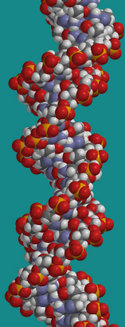 Дезоксирибонуклеиновата киселина (ДНК) е нуклеинова киселина, която носи генетичните инструкции за биологическото развитие на всички клетъчни форми на живот и много от вирусите. ДНК понякога се нарича молекула на наследствеността, тъй като тя се наследява и се използва за разпространение на признаци. При възпроизводството тя се реплицира и се предава на потомството.
Дезоксирибонуклеиновата киселина (ДНК) е нуклеинова киселина, която носи генетичните инструкции за биологическото развитие на всички клетъчни форми на живот и много от вирусите. ДНК понякога се нарича молекула на наследствеността, тъй като тя се наследява и се използва за разпространение на признаци. При възпроизводството тя се реплицира и се предава на потомството.
При бактериите и други прости клетъчни организми ДНК е разположена някъде из цялата клетка. В сложните клетки, от които са изградени растенията, животните и в други многоклетъчни организми, повечето ДНК ев хромозомите, които се намират в клетъчното ядро. Органелите–енергийни генератори, познати като хлоропласти и митохондрии също носят ДНК, както и много вируси.
* Гените могат да се разглеждат грубо като готварска книга на организма.
* Една верига ДНК съдържа гени – области, които регулират гените – и други области, които нямат функции или функциите им все още не са познати;
* ДНК се състои от две нишки с връзки помежду им, които могат да се разделят подобно на цип;
* ДНК кодира генетичната информация благодарение на четири „строителни елемента”, наречени бази: аденин, тимин, гуанин, цитозин. Те се обозначават съкратено като А, Т, Г и Ц и имат свойството всяка да „се чифтосва” само с една от останалите три бази: А Т, Т А, Г Ц, Ц Г; такаче едно „А” на едната нишка от ДНК ще се „съеши” успешно само с „Т” от другата нишка.
* Редът е от значение: А Т не е еквивалентно на Т А, както и Ц Г не е едно и също с Г Ц.
* Но, тъй като са възможни само 4 комбинации, базите на едната от нишките са достатъчни, за да се опише последователността.
* Редът, в който са разположени базите по дължината на ДНК, е важен —ДНК-последователността (или секвенцията) е описанието на гените.
* Репликацията или удвояването на ДНК се осъществява чрез разделяне на двете нишки с относително прости химически реакции и създаване на„втората половина” на така получената единична верига чрез потапяне в„супа”, която съдържа всичките четири бази. Тъй като всяка база може дасе комбинира само с една от останалите три бази, подредбата на базите в съществуващата верига определят еднозначно какви бази ще има в новообразуваната верига и как ще са подредени. По този начин, всяка единична верига образува точно копие на оригиналната ДНК, като събира необходимите бази в „супата”, освен ако не настъпи мутация.
* Мутациите са просто химически грешки в този процес: една база може случайно да бъде пропусната, вмъкната или грешно копирана или пък веригата може да бъде скъсена или удължена; всички други основни мутации могат да се опишат като комбинация от тези случайни „операции”.
ДНК е позната на учените от повече от сто години. Първоначално е идентифицирана през 1868 г. от швейцарския биолог Фридрих Мишер в сперматозоиди от сьомга. Той нарича откритата от него субстанция нуклеин, но въпреки това ДНК е призната официално едва през 1943 г. в резултат на експеримент, реализиран от Осуалд Ейвъри. Структурата на двойната ДНК спирала е била открита през 1953 г. от Джеймс Уотсън и Франсис Крик в Кеймбриджкия университет. За това си откритие те получават Нобелова награда за химия.
http://bgnauka.eu/pic/bio/DNA1.jpg
Въпреки че понякога ДНК е наричана „молекулата на наследствеността”, парчетата ДНК, както си ги представят повечето хора, не са единични молекули. По-скоро те са двойки молекули, които се преплитат като лиани,образувайки двойна спирала.
Всяка лианообразна молекула е ДНК верига: химически свързана верига отнуклеотиди, всеки от които се състои от захар, фосфат и един отчетирите вида ароматно-въглеводородни „бази”. Тъй като нишките в ДНА са изградени от тези нуклеотидни елементи, те са полимери.
Тъй като има четири вида бази, има и четири вида нуклеотиди, които често се обозначават като своите бази. Тези бази са аденин (А), тимин (Т), цитозин (Ц) и гуанин (Г).
Полинуклеотидните вериги в двойната верига на ДНК могат да се свързват чрез хидрофобния ефект. Това, кои вериги остават свързани, се определя от комплементарното свързване. Всяка база образува водородни връзки само с една от останалите бази — А с Т, Ц с Г — така че видът на базитена едната верига определя силата на връзката — колкото повече комплементарни бази съществуват, толкова по-силна и устойчива е връзката.
Механизмите в клетката могат да разделят двойната спирала на ДНК и такавсяка единична верига може да служи за шаблон за създаване на нова верига, почти еднаква с предишната. Грешките при синтеза се наричат мутации. Процес, наречен ПВР имитира този процес ин витро в не жива среда.
Тъй като при свързване в двойки нуклеотидните бази се обръщат към остана спиралата, а захарните и фосфатните групи са от външната страна,двете вериги, които те образуват, изглеждат като носещи елементи на спиралата. В действителност химическите връзки между фосфатите и захарите свързват един нуклеотид със съседните му във веригата.
Последователността от нуклеотиди по една ДНК-верига в един ген дефинира един белтък, който организмът трябва да бъде произведе или както е прието да се казва, да „експресира” в един или няколко момента отживота си, като използва информацията, съдържаща се в последователността. Отношението между нуклеотидната последователност и тази от аминокиселини в белтъка се определя от прости клетъчни правилана транслация, познати под името генетичен код. Генетичният код се състои от трибуквени думи (наречени кодони), образувани от последователност от три нуклеотида (напр. АЦТ, ЦАГ, ТТТ). Тези кодони след това могат да се транслират посредством информационна рибонуклеинова киселина (РНК) и впоследствие транспортна РНК до кодон,съответстващ на определена аминокиселина. Тъй като има 64 възможни кодона, повечето аминикиселини имат повече от един възможен съответе нкодон. Има също така три „стоп” или „безсмислени” кодо на, озачаващи края на кодовата последователност.
Изглежда, че при много видове организми само малка част от цялата последователност на геномa кодира белтъци. Функцията на останалата час те предмет на спекулации. Знае се, че някои нуклеотидни секвенции задават афинитет към ДНК свързващи белтъци, които играят различни жизненоважни роли, по-специално чрез контрол на репликацията и транскрипцията. Тези последователности често се наричат регулационни секвенции и изследователите предполагат, че засега са идентифицирали само една малка част от съществуващите. „Отпадъчната ДНК” представлява участъци, които не изглежда да съдържат гени или да имат някаква функция.
Секвенцията определя и податливостта на даден сегмент от ДНК на разкъсване, причинено специфично от ензими – рестрикционни ендонуклеази (съкр. рестриктази)— най-типичното средство на генното инженерство.Позициите на местата на разкъсване след третиране на генома на даден индивид с определен ензим са строго специфични. В практиката се използва третиране само на определен участък, при което може да се състави ДНК профил на индивида (един от множеството възможни).
Репликация или синтез на ДНК е процесът на копиране на двойната верига на ДНК преди деление на клетката. Двете получени двойни вериги са почти съвсем еднакви, но понякога грешки при репликацията могат да доведат дополучаване на не съвсем точно копие (виж мутация) и всяка от тях се състои от една оригинална и една ново синтезирана верига. Това се нарича полуконсервативна репликация. Процесът на репликация се състои от три стъпки: инициация, репликация и терминация.
Водородните връзки между веригите на двойната спирала са достатъчно слаби, за да могат лесно да бъдат прекъснати с помощта на ензими. Ензими, познати като хеликази разсукват веригите, за да подпомогнат действието на ензими, четящи последователността като ДНК-полимеразата. При разсукването ензимите трябва да прекъснат по химически на чинфосфатният гръбнак на едната от веригите, за да може тя да се завърти около другата верига. Веригите могат също така да бъдат разделени чрез леко нагряване, използвано например в ПВР, ако имат по-малко от около10 000 базисни двойки (10 кило базисни двойки или 10 кбд). Усукването на ДНК-веригите затруднява разделянето на дълги сегменти.
Когато двата края на една двойна спирала на ДНК са съединени, така че тя образува кръг, както е при плазмидите, веригите са топологично заплетена. Това означава, че те не могат да бъдат разделени чрез леко нагряване или друг процес, който не включва прекъсване на едната верига. Задачата за разплитане на топологично свързани вериги на ДНК се изпълнява от ензими, наречени топоизомерази. Някои от тези ензими разплитат цикличната ДНК чрез прекъсване на двете вериги, така че друг двойно верижен сегмент да може да мине между тях. Разплитането е необходимо за репликацията на циклични ДНК, както и за различни видове рекомбинация в линейни ДНК.
Спиралата на ДНК може да приеме една от три леко различаващи се форми.За преобладаваща в клетките се счита „B” формата, описана от Джеймс Уотсън и Франсис Крик. Тя е широка 2 на нометра и има дължина 3,4нанометра на 10 базисни двойки (бд) от последователността. Средната дължина на стъпката на спиралата — последователността, в продължение на която спиралата прави едно пълно завъртане около оста си – е също толкова. Честотата на завъртанията зависи до голяма степен от притеглящите сили, които всяка база упражнява върху съседите си във веригата.
Малката ширина на двойната спирала прави намирането й чрез конвенционална електронна микроскопия невъзможно, освен чрез интензивно оцветяване. В същото време ДНК в много клетки може да бъде с макроскопична дължина — около 5 сантиметра за веригите в една човешка хромозома. Следователно клетките трябва да сбиват или „пакетират” ДНК,за да могат да я побират. Това е една от функциите на хромозомите,които съдържат белтъци, подобни на макари, наречени хистони, околокоито ДНК се навива.
При липса на напрежение B-формата на ДНК-спиралата се усуква на 360° на всеки 10,6 базови двойки. Но много процеси в молекулярната биология могат да причинят напрежения. Един сегмент от ДНК с недостатъчно или прекалено усукване на спиралата се нарича съответно положително или отрицателно „пренавит”. ДНК ин виво обикновено е отрицателно пренавита,което подпомага разсукването на двойната спирала, необходимо за РНКтранскрипция.
Другите две познати двойно-спирални форми на ДНК, наречени A и Z, се различават донякъде по своята геометрия и размери. Изглежда възможно A-формата да се среща само в дехидрирани проби от ДНК като тези,използвани в кристалографски експерименти, и може би в хибридни свързвания на ДНК и РНК-вериги. ДНК-сегменти, които клетките са метилирали с регулиращи цели, могат да възприемат Z-геометрия, при която веригите са усукани около оста на спиралата като огледален образна B-формата.
Поради несиметричната форма и свързване на нуклеотидите една ДНК-верига винаги има видима ориентация. Поради тази ориентация внимателното разглеждане на една двойна спирала показва, че нуклеотидите са ориентирани в една посока по едната верига („възходящата верига”) и в обратната посока по другата верига („низходящата верига”). Такова разполагане на веригите се нарича анти паралелно.
Заради химическата номенклатура хората, които работят с ДНК, наричат асиметричните краища на едната верига 5′ и 3′ краища(произнасят се „пет прим” и „три прим”). Работещите с ДНК, (а също така и ензимите), винаги четат нуклеотидните последователности в „5′ към 3′ посока”. В една вертикално ориентирана двойна спирала 3′ веригата се нарича възходяща, а 5′ веригата — низходяща.
Като резултат от тяхното анти паралелно разположение и предпочитанията на ензимите при четене на последователността, дори и двете вериги да носеха идентични, вместо допълващи се поредици, клетките могат датранслират правилно само едната от тях. Другата верига може да бъдепрочетена от клетката само отзад напред. Молекулярните биолози наричат една последователност „сенс”,ако е транслирана или транслируема, а нейната допълнителна (комплементарна) — „антисенс”. Следователно, донякъде парадоксално,шаблонът за транскрипция е антисенс-веригата. Полученият запис е РНК-копие на сенс-веригата и самият той е сенс.
При някои вируси разликата между сенс и антисенс е размита, тъй като определени секвенции от техните геноми вършат двойна работа, кодирайки един белтък, ако бъдат четени 5′ към 3′ по едната верига и втори белтък, когато се четат в обратна посока по другата верига. Така геномите на тези вируси са необикновено компактни за броя гени, които съдържат. Това се разглежда от биолозите като адаптация.
Тополозите обичат да отбелязват, че разполагането на 3′-края на едната верига на ДНК до 5′-края на другата в двата края на един двойно спираленсегмент прави конструкцията „раков канон”.
В някои вируси ДНК е в неспирална, едноверижна форма. Тъй като много от механизмите за поправка на ДНК в клетките работят само със свързани бази, вирусите, които носят геномите си в едно верижна ДНК, мутиратсравнително по-често. В резултат такива видове могат да се адаптират по-бързо, за да не изчезнат като вид. Резултатът обаче не би бил толкова благоприятен при по-сложни и по-бавно размножаващи се организми, което може да обясни защо само вирусите носят едноверижни ДНК. Тези вируси печелят и от по-ниската „цена“ на репликацията на една верига вместо две.
")



 и усойница (Vipera berus)")
